Asexual Reproduction (Experiment):
Introduction:
Annelids are well renown for their ability to regenerate lost parts. Some annelids are able to regenerate both anterior and posterior ends from just a single segment. Whilst most annelids seem to possess the ability to regenerate their posterior end the ability to regenerate the anterior is less universal (Bely 2006). Annelids able to successfully regrow both ends are usually smaller species although some larger members from the Amphinomidae family have demonstrated this ability as well. This regenerative ability allows for asexual reproduction to occur via fragmentation and/or fission (Ahrens et al. 2014).
Studies have been undertaken on the regenerative ability of Hermodice carunculata and Eurythoe complanata yet none have been done on Eurythoe laevisetis (Ahrens et al. 2014). Eurythoe laevisetis is a close relative of Eurythoe complanata with the only difference in external morphology being that Eurythoe complanata has three types of notochaetae present and Eurythoe laevisetis only has two (Barroso et al. 2010). Whilst it is expected that Eurythoe laevisetis will possess the ability to regenerate both posterior and anterior ends as well, a formal experiment will be run on it’s regenerative ability to confirm this.
Methods:
Specimens were collected off the Heron Island Reef and brought back to the aquarium. Originally housed together, once experimentation began they were placed into separate clear plastic containers that had a mesh lid. Each container had inside pieces of the rock, which the fireworms were originally extracted from, to provide housing. Other organisms, both dead and alive, such as crabs, shrimp, peanut worms and brittlestars were collected off the same rocks and placed into the containers with the fireworms to act as a food source.

Figure 8: The plastic container that the fireworms were housed in.
Catherine Russo, University of Queensland St. Lucia, 2014
During Week 1 of the experiment only one (Worm1) Eurythoe laevisetis was amputated. This was completed by pulling on one half of the worm with a pair of tweezers whilst the other half remained inside a tube. Once the worm was split in half the tube was delicately cracked open and the rest of the fireworm retrieved. Both halves of the worm were then placed within the same container and left in the aquarium for a week.
During Week 2 of the experiment the remaining two (Worm2 & Worm3) Eurythoe laevisetis were amputated. These worms were pulled out of the water one at a time and placed on a folded piece of paper towel. The worms were then measured and the total length of each worm was divided by three. Once the appropriate measurement was determined to correctly cut the worms into thirds, each worm was cut using a scalpel to make the incision and tweezers to prevent unnecessary movement of the worm. All sections belonging to the same worm were placed into the same container.
The worm that was split in half in Week 1 was examined under a compound microscope. Observations were recorded. During Week 3 of the experiment all of the worm segments were taken out of their containers and examined under a microscope. Observations were recorded.
Results:
Worm1 segments displayed regrowth of both the anterior and posterior end. Whilst the newly grown posterior end did not change much in appearance over the course of two weeks the newly grown anterior end did. Development and regrowth of a new head could clearly be seen.

Figure 9: Formation of a new head one week after amputation (Worm1).
Catherine Russo, University of Queensland St. Lucia, 2014
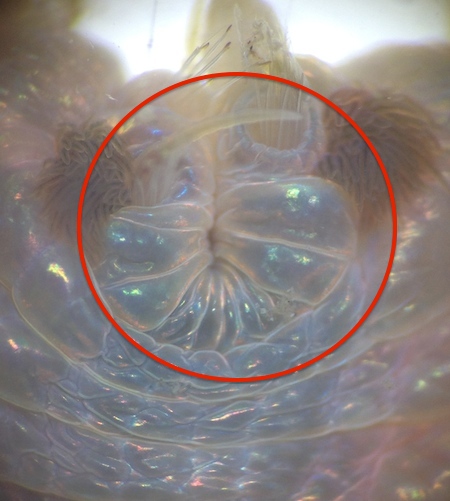
Figure 10: Formation of a new posterior end one week after amputation (Worm1).
Catherine Russo, University of Queensland St. Lucia, 2014
All of the segments belonging to all of the worms were alive at the end of the experiment. Whilst Worm2 & Worm3 segments were not at developed as the Worm1 segments, each segment showed the growth of a new anterior and/or posterior end.
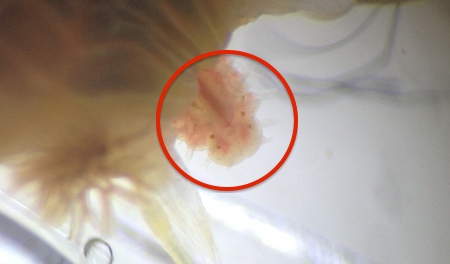
Figure 11: Formation of a new head two weeks after amputation (Worm1).
Catherine Russo, University of Queensland St. Lucia, 2014

Figure 12: Five of the fireworm fragments clustered together on one side of a petrie dish.
Catherine Russo, University of Queensland St. Lucia, 2014
Discussion:
This study has shown that the posterior and anterior ends of Eurythoe laevisetis can be regenerated both individually in a segment that has only lost one end and simultaneously in a segment that has lost both. This information does not come as a surprise due to the number of species capable of asexual reproduction by fragmentation in the Amphinomidae family. In fact, the ability of regeneration is so widespread that Bely (2006) argues that regeneration is an ancestral feature of the Annelida and has probably been lost repeatedly over time due to evolution.
In Euythoe complanata regeneration occurs in two steps. The first step involves the primary neuronal structures forming around the ‘old’ ventral nerve cord. The second step is where new neurons are formed in the regenerated area that are able to reconnect with the ‘old’ nerve cord by utilizing their outgrowing axons (Muller et al. 2003). Whilst studies on Eurythoe laevisetis would have to be completed to confirm that this process is the same, since the two species are so similar it is assumed that the process is consistent between both species.
To further this experiment a larger sample size and a longer time frame to determine how long it takes for the anterior end to completely regrow would be beneficial. A more in depth look at how both ends actually regrow, to see if it is the same as Euythoe complanata, and whether or not certain factors like temperature or polluted water affect regrowth rates would also be advantageous. |